第五章 DNA甲基化
5.1 DNA甲基化概况
-
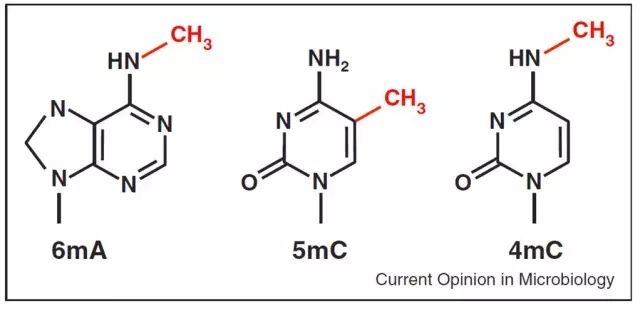
1.在真核生物中,甲基化只发生在胞嘧啶第5位的碳原子上,是由DNA甲基转移酶所催化,以S-腺苷甲硫氨酸(SAM)作为甲基供体,将甲基转移到胞嘧啶上,生成5-甲基胞嘧啶的一种反应。
-
2.在哺乳动物中,DNA甲基化主要发生在CpG双核苷酸序列的胞嘧啶上。
-
3.根据作用方式和参与反应的酶的不同,甲基化反应可以分为:
维持甲基化(maintenance methylation):
与DNA复制相关联,当甲基化的双链DNA被复制生成两条新的双链DNA以后,只有亲代链是甲基化的,而新合成的子代链是非甲基化的。DNA甲基化转移酶(DNMT1)以非对称甲基化DNA为底物,识别新生成的DNA双链中亲代单链上已经甲基化的CpG位点,然后催化互补单链相应的位置的胞嘧啶(C)发生甲基化,以维持新生成的双链DNA的甲基化。
从头甲基化(de novo methylation):
是对DNA上甲基状态的重新构建,它不依赖DNA复制,在完全非甲基化的DNA碱基位点上引入甲基,是甲基化的建立机制。
-
4.一般认为,维持甲基化的主要参与酶是DNMT1,从头甲基化则依赖于DNMT3a和DNMT3b的活性。
-
5.DNA甲基化可能存在于所有高等生物中,基因组中60% ~ 90%的CG序列都存在甲基化现象,但甲基化的DNA在整个基因组中所占比例通常很小。
5.3 真核生物DNA甲基化修饰系统
真核生物细胞中甲基化模式的建立和维持依靠甲基化修饰系统来完成,该系统中包含两类重要蛋白质:DNA甲基转移酶(DNMTs)和甲基结合蛋白(MBDs)。
5.4 DNA甲基化与遗传物质的稳定性
DNA甲基化作为一种标签决定了复制起点与细胞膜的结合,控制了复制起始,使得DNA复制与细胞分裂保持一致。
5.4.2 DNA甲基化与错配修复
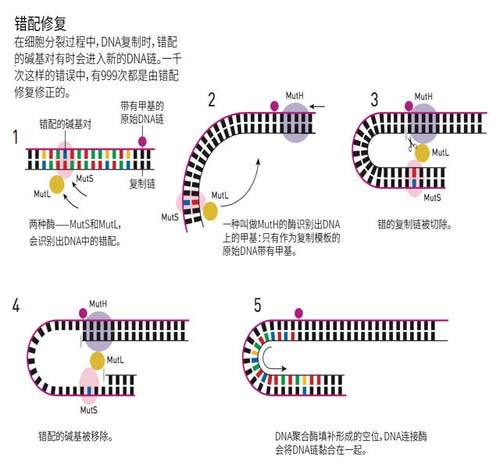
由于复制时母链上的5’-GATC-3’中的腺嘌呤第6位N是甲基化的,而子链上的5’-GATC-3’是非甲基化的,在合成后2-5min才会被DNA甲基化酶甲基化。因此复制后双链DNA在数分钟内保持半甲基化状态,错配修复系统会依据“保持母链,修复子链”的原则,对参入错误的碱基进行修复,保证复制的忠实性。
5.4.3 DNA甲基化与转座子失活
在人类基因组中,约1/3的序列为散乱的内源性转座子DNA,细胞中90%的甲基化位于转座子中,呈高度甲基化的转录沉默状态。
5.5 DNA甲基化与基因表达调控
DNA甲基化在转录水平调控基因的表达,尤其是转录起始阶段。DNA甲基化对基因的抑制活性是多方面的作用导致的,DNA甲基化可能直接影响一些转录因子的结合活性,或DNA甲基化结合蛋白抑制了转录因子的结合。
5.5.1 DNA甲基化直接影响一些转录因子的结合活性
- 在细胞中有些转录因子的特异性结合位点中有CpG位点,当这些位点发生甲基化的时候,就降低了转录因子与启动子结合的效率,从而降低了基因的转录率;
- DNA的大沟内,是众多转录因子与DNA结合的部位,由胞嘧啶甲基化生成的5-甲基化胞嘧啶会伸入DNA双螺旋的大沟,从而影响转录因子的结合。
5.5.2 DNA甲基化结合蛋白与转录抑制
MBD家族蛋白都含有两类结构域:一类是MBD结构域,可以与甲基化或半甲基化的DNA相互结合;另一类是可以与多种转录抑制因子相互结合以发挥抑制基因表达活性的作用。
5.6 DNA甲基化与其他表观遗传修饰的关系
- DNA甲基化、组蛋白甲基化、染色质的紧密程度:与DNA的不可接近性以及基因处于抑制和静息状态相关;
- DNA的去甲基化、组蛋白的乙酰化、染色质压缩状态的开启:与转录的启动、基因活性相关。
5.6.1 DNA甲基化与核小体定位
DNA甲基化修饰程度与核小体序列中有相似的10bp周期性,并且核小体占据区甲基化的程度明显高于核小体缺乏区,这表明,基因组核小体定位强烈影响甲基化模式,DNA甲基转移酶倾向于把核小体序列作为靶序列。
5.6.2 DNA甲基化与组蛋白修饰
- 组蛋白的甲基化和乙酰化修饰对染色体活性的调节很重要;
- 组蛋白的甲基化对DNA的甲基化有一定的影响。
(1)组蛋白H3K9三甲基化可以指导DNA甲基化,为DNA甲基化的必要条件(e.g. dim-5突变体会导致基因组低甲基化,dim-5为H3K9甲基化酶);
(2)组蛋白甲基化引起DNA甲基化:引起H3K27甲基化的复合物EZH2可与DNA甲基转移酶结合,引起相应位点DNA的甲基化。
(3)识别甲基化CpG位点的MBD蛋白可以与HDAC家族相互作用,从而抑制基因的表达;
(4)甲基转移酶也可与HDAC相互作用;
(5)组蛋白乙酰化的改变可以影响DNA甲基化的状态。
5.6.3 DNA甲基化与非编码RNA
非编码RNA可以通过调节DNMT的表达进而调节DNA的甲基化。
第六章 RNA可变剪接的表观遗传学机制
6.1 引言
基因组携带有两类遗传信息:
- 一类提供生命必须的蛋白质的模板,称为遗传编码信息;
- 另一类提供基因选择性表达(何时、何地、何种方式)的指令,称为表观遗传信息。
6.2.1 可变剪接的基本概念
-
哺乳动物RNA由长度约140核苷酸的外显子及中间穿插长度约上千核苷酸的非编码内含子组成;
-
RNA剪接:是指依据RNA顺式和反式元件,在包括表观遗传信息在内的剪接指令指导下,剪接体正确识别、连接外显子并精确去除长内含子,从而加工为成熟mRNA的过程。
-
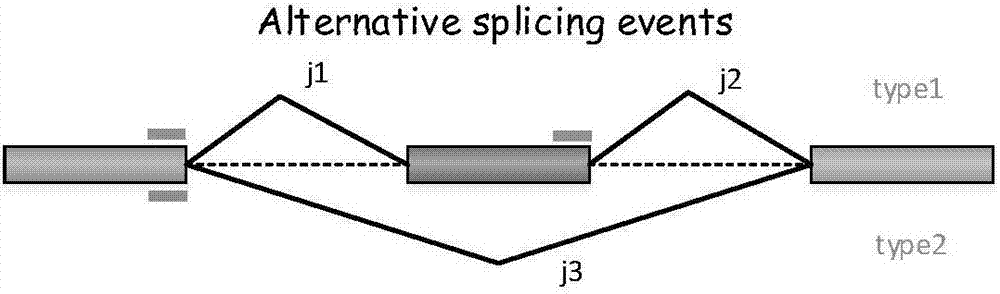
可变剪接:是指从一个mRNA前体通过不同的剪接方式(即选择不同的剪接位点组合)产生不同的mRNA剪接异构体的过程;
-
mRNA前体的可变剪接是真核生物基因表达调控的重要方式和产生蛋白质多样性的重要机制。
6.2.2 可变剪接的基本类型
- 第一类是外显子跳跃,中间盒式外显子与两侧内含子序列一起被剪去;
- 第二、三类是可变3’、5’剪接位点;
- 第四类是内含子保留(植物、真菌普遍);
- 第五类是互斥外显子;
- 第六类是第一外显子可变;
- 第七类是最后外显子可变。

6.5 可变剪接的表观遗传学机制
剪接与转录耦合指出调控转录的因子也可调控剪接。事实上,剪接位点的选择远比想象中的复杂。RNA结合元件、RNA聚合酶Ⅱ延伸速率均不足以解释可变剪接的精巧机制。
第十一章 表观遗传学的功能
干细胞的分化、X染色体失活、基因组印记等等。
11.3 基因组印记
基因组印记涉及DNA甲基化以及位点特异性的、差别的染色质结构。几乎每一个印记基因中都含有一个或多个差别的甲基化区域。DNA甲基化在基因组印记擦除和重建周期中起着至关重要的作用。
除了DNA甲基化,一些印记基因还需要组蛋白的修饰。
补充:
- 通常,在二倍体动物中,来自于父本和母本的两个等位基因在胚胎和动物中都会表达。然而,一些基因被特异性地印记化,以至于仅仅在父本或母本的基因是有活性的。因此,虽然胚胎从其双亲那里接受了相同的遗传信息,然而,双亲的基因组在功能上并不完全相同。